


Dugo poznata biljka s novim perspektivama
Ekstrakti Urtica dioica imaju terapeutski potencijal za ublažavanje oksidativnog stresa i hiperglikemije izazvane dijabetesom.
Kopriva Urtica dioica L., s malom koprivom Urtica urens, predstavljaju najzastupljeniju vrstu roda Urtica koji obuhvaća 63 vrste cvjetnica i pripada porodici Urticaceae (40 rodova i više od 500 vrsta) glavnih Skupina kritosjemenjača.
14. sijećanj 2022.
Kopriva, dugo poznata Vlaknasta biljka s novim perspektivama
Sažetak
Kopriva Urtica dioica L. je višegodišnja kultura s malim zahtjevima, dobro prilagođena širokom rasponu uvjeta okoliša. Uspješno se uzgaja u većini europskih klimatskih zona, istovremeno promičući raznolikost lokalne flore i faune. Uzgoj koprive mogao bi pomoći u zadovoljavanju snažnog porasta potražnje za sirovinama na bazi biljnih vlakana kao zamjene za umjetna vlakna u sektorima tako raznolikim kao što su tekstilna i automobilska industrija. U ovom pregledu predstavljamo povijesnu perspektivu selekcije, berbe i značajki prerade vlakana gdje je detaljno prikazano stanje tehnike odabira sorti koprive. Omogućena je sinteza općeg znanja o njegovoj biologiji, prilagodljivosti i genetičkim sastavnim dijelovima, naglašavajući nedostatke u našem trenutnom znanju o interakcijama s drugim organizmima. Nadalje smo se pozabavili značajkama uzgoja i prerade, stavljajući poseban naglasak na sustave žetve i procese ekstrakcije vlakana kako bismo poboljšali prinos i kvalitetu vlakana. Opisane su različite upotrebe u industrijskim procesima, a posebno za obnovu rubnih zemljišta i putevi budućih istraživanja ove višenamjenske biljke visoke vrijednosti za globalno tržište vlakana.
Ključne riječi:
Urticadioica L., kopriva, filogenija, uzgoj, proizvodnja i prerada vlakana, fitomenadžment
1. Uvod
Urtica dioica L. naziva se “velika kopriva”, ali je kolokvijalno iu literaturi poznata samo kao “kopriva” [1,2,3,4]. Kopriva Urtica dioica L., s malom koprivom Urtica urens, predstavljaju najzastupljeniju vrstu roda Urtica koji obuhvaća 63 vrste cvjetnica i pripada porodici Urticaceae (40 rodova i više od 500 vrsta) glavnih Skupina kritosjemenjača. Ove dvije vrste porijeklom su iz Europe, Afrike, Azije i Sjeverne Amerike [5]. Urtica dioica L. prvi put je opisao Carl von Linné 1753. godine. Ime „Kopriva“ moglo bi potjecati od anglosaksonske riječi „noedl“ što znači igla, a Urtica je latinska riječ koja znači „paliti“, što se odnosi na žarenje izazvano dodirivanjem biljke. “Dioica” se odnosi na činjenicu da se muški i ženski cvjetovi nalaze na odvojenim biljkama [3]. Kopriva (Urtica dioica) koristila se kao hrana i vlakna još od srednjeg vijeka. Uz lan i konoplju, kopriva je bila najvažniji biljni tekstilni materijal u Europi jer za razliku od pamuka raste čak iu sjevernim podnebljima. Njemačka i Austrija bile su pioniri u uzgoju koprive tijekom 19. stoljeća i započele su komercijalni uzgoj vlakana koprive (Slika 1). Uz sankcije nametnute pamuku tijekom Prvog svjetskog rata, njemačka vojska koristila je tkaninu od koprive za uniforme svojih vojnika. Međutim, jeftinija vlakna iz jednogodišnjih usjeva bila su lakše dostupna nakon Drugog svjetskog rata [1]. Nedavno je sve veća zabrinutost oko upotrebe neobnovljivih izvora u proizvodnji dovela do obnovljenih interesa za ponovnu upotrebu prirodnih ili bioloških vlakana [6] (Slika 1). Studije predviđanja predviđaju daljnji snažan razvoj tržišta za vlakna dobivena iz biljaka, s procijenjenim povećanjem od 300% u sljedećih 25 godina [7]. Predviđeno je da bi površina zemljišta potrebna za biljna vlakna za materijalne potrebe mogla doseći čak 300 000 ha do 2035. [8]. U Europi su glavne biljke koje se koriste uglavnom lan i konoplja, s relativnim tržišnim udjelom na tržištu biovlakana od 64% odnosno 10% [7]. Međutim, postoje dobri ekonomski i ekološki razlozi za uzgoj Urtice dioice kao predivačke kulture: (i) to je višegodišnji usjev s malim zahtjevima za gnojivima i pesticidima [9], (ii) postoji visok potencijal uzgoja u nekoliko područja koja omogućiti regionalnu proizvodnju [10], (iii) može poboljšati tla preopterećena nitratima i fosfatima, jer je kopriva nitrofilna zeljasta biljka [9], (iv) promiče raznolikost lokalne flore i faune [10], i (v) može se proizvoditi na zemljištu nepogodnom za proizvodnju hrane, uključujući onečišćena zemljišta. Proizvodnja uzgojenih usjeva na marginalnim zemljištima koja su neprikladna za proizvodnju hrane mogla bi pomoći u ublažavanju potencijalnih sukoba između proizvodnje hrane i neprehrambenih proizvoda. Kopriva ima još jednu kartu u ovom kontekstu, budući da snažno raste posvuda, bez intenzivnih unosa kao što su pesticidi, herbicidi ili navodnjavanje, čak i u prilično siromašnom tlu [11]. Kopriva također raste u hladnim klimatskim područjima [12], što je čini relevantnom biljkom kandidatom za lokalnu proizvodnju i preradu diljem Europe. Urtica dioica također se često povezuje s topolama [11] i vrbama u obalnim staništima diljem Europe [13], koje se također intenzivno koriste u praksama fitomenadžmenta. 00
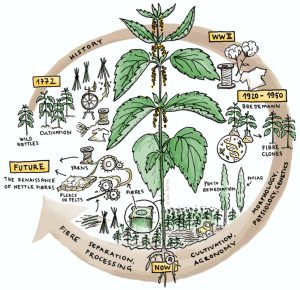
Slika 1 Grafički prikaz povijesnog napretka u razvoju koprive.
Ovaj pregled pruža sveobuhvatnu obradu prilika koje se pojavljuju u proizvodnji vlakana koprive, posebno u vezi s prenamjenom marginalnih zemljišta i proizvodnjom obnovljivih izvora vlakana. Podijeljen je u četiri odjeljka prikazana na slici 1, koji opisuju (i) povijest upotrebe koprive (ii) biologiju, fiziologiju i genetiku Urtica dioica L., (iii) uzgoj, žetvu i preradu vlakana i ( iv) različite moguće lance vrijednosti i uporabe koprive. Cilj mu je sažeti dostupno znanje o korištenju koprive za vlakna kako bi se pružila osnova za razumijevanje njezine primjene, te također istaknuti gdje bi poboljšanja u proizvodnji usjeva i razvoju industrijskih vlakana bila korisna. Identificirali smo nova polja u usponu u ovom istraživačkom području, privlačeći, nadamo se, nove istraživače.
2. Povijesna perspektiva
Kopriva je stoljećima dio ljudskog društva i naširoko je cijenjena zbog svog naizgled beskonačnog niza upotrebe. Budući da kopriva ponovno dolazi u središte pozornosti, u ovoj se rubrici osvrćemo na povijest njezina odabira, berbe i prerade vlakana.
2.1. Odabir klonova
Kopriva (Urtica dioica L.) se stotinama godina prerađivala u tekstil. Sačuvano tkivo koprive dokazuje njezinu upotrebu u Švicarskoj još u 7. stoljeću [14]. Prva pisana veza između “koprive” i “Urtice” može se datirati u 1391. godinu u Velikoj Britaniji [15]. Prvi industrijski pokušaji korištenja stabljike koprive za opskrbu tekstilnih vlakana datiraju iz 1850. godine i sljedećih desetljeća [16,17]. Stoga ima dugu povijest kao biljka vlakana u Njemačkoj i Austriji gdje se koristio, zajedno s lanom (Linum usitatissimim L.) ili konopljom (Cannabis sativa L.) za tekstil prije pojave pamuka (roda Gossypium) [18]. ]. Mnogi klonovi koprive koji se trenutno razvijaju za primjenu vlakana potječu iz kolekcija koje su inicijalno prikupljene tijekom tog razdoblja. Gustav Bredemann, njemački poljoprivredni znanstvenik i botaničar, i drugi započeli su 1920-ih sa zbirkom izvorno samoniklih biljaka koprive i odabirom obećavajućih primjeraka u pogledu njihove snage i sadržaja vlakana [17]. Vlaknaste koprive iz ove selekcije uzgojene su i pokazale su sadržaj vlakana do 17,6% (sadržaj čistih vlakana nakon kemijske separacije), najmanje tri puta veći od udjela divljih kopriva [17]. Nakon 1945. interes se smanjio, iako je oko 30 kultivara, odnosno klonova (sortiman hamburške koprive), održavano i čuvano od strane Instituta za primijenjenu botaniku Sveučilišta u Hamburgu [19]. Godine 1991., Poljoprivredni institut pokrajine Thüringen” (TLL—Thüringer Landesanstalt für Landwirtschaft), započeo je istraživanje dodatnog niza klonova koprive koje je održavao bivši Savezni istraživački centar za poljoprivredu (FAL, Braunschweig, Njemačka) [19] . Prinosi biomase do 90 dt ha−1 i sadržaj vlakana (sadržaj čistih vlakana) nakon kemijske separacije do 14% pronađeni su tijekom razdoblja od četiri godine, na temelju metode procjene iz 1942. [20]. Ti su klonovi ponovno procijenjeni 1993. godine. U početku je rad bio usmjeren na istraživanje razvoja biljaka, prinosa i fenotipa biljnog materijala jer je većina izvornih opisa i podataka o učinkovitosti izgubljena tijekom vremena [21]. Godine 1993. Dreyer je započeo s ovim rasponom biljnog materijala u 4-godišnjem ciljanom multifaktorijalnom eksperimentu uzgoja (klon, gnojidba) s, uz ostalo, određivanjem fenotipske varijabilnosti, karakteristika lišća i stabljike kao što su prinos biomase i sadržaj vlakana . Od testiranih klonova, utvrđeno je da “klon B13” daje najbolje prinose vlakana i suhe mase stabljike [17] i da se najviše koristio u eksperimentalnim poljima [2,21]. Francken-Welz izvješćuje o poljskim pokusima na pjeskovitoj ilovači u blizini Bonna (zapadno od Njemačke) 1997. Osnova koprive je Thuringijski sortiment, što ne dopušta nikakvu jasnu referencu na izvornu Bredemannovu klonsku osnovu. Rezultati istraživanja prinosa utvrđeni su nakon 2 i 3 godine uzgoja. Zabilježen je prinos biomase od 65 do 82 dt ha−1 sa srednjim sadržajem vlakana od 17,3% pri gustoći sadnje koja varira od 1,7 do 5 biljaka m−2 i srednjoj stopi gnojidbe dušikom od 140 kg ha−1 [22]. Može se pretpostaviti da je sadržaj vlakana određen laboratorijskom mehaničkom dekortikacijom. Dodatni rezultat pokusa je dokumentacija o mogućem uzgoju koprive na bazi sjemena. Ova metoda rezultira povećanim naporom i rizikom, kao i manjom konkurentnošću na početku uspostavljanja usjeva. Ovi rezultati potkrijepljeni su eksperimentom iz Thuringijskog rada [23].
Istraživački projekt usmjeren na uzgoj financirala je Njemačka savezna zaklada za okoliš (DBU—Deutsche Bundesstiftung Umwelt, Osnabrück, Njemačka) od 2008. do 2012. Glavni cilj bio je razviti štedljiv postupak razmnožavanja koprive na temelju somatske embriogeneze i stvaranje inkapsuliranih sintetičkih sjemenki [24]. Osim stvaranja brojnih temeljnih znanja o koracima procesa, projekt nije bio uspješan, uglavnom zbog fitosanitarnih razloga u tijeku nastanka i razvoja kalusa. Nadalje, poseban napor uložen je u uzgoj novih klonova s višim sadržajem vlakana na temelju originalnog asortimana hamburgera Bredemanna u istovremenom projektu koji je financiralo Savezno ministarstvo hrane i poljoprivrede, Berlin, Njemačka. Tri godine i dvije lokacije (Hannover i Soltau, Donja Saska, Njemačka) pokusi s odabranim Bredemannovim i novim uzgojenim klonovima pokazali su visoku varijabilnost fenotipskih karakteristika i karakteristika povezanih s prinosom. Konkretno, oplemenjivanje je stoga teže jer je za pouzdanu selekciju potrebna odgovarajuća baza podataka iz dugogodišnjeg uzgoja u polju [25]. Polazna točka rada bilo je šest izvornih klonova Bredemanna i četiri druga koja su već uzgojena (“Z”) od njih. Odabir od osam novih kultura (od kojih pet ima Bredemannov klon B 13 u generaciji roditelja) okarakteriziran je kao obećavajući za daljnje razmnožavanje i testiranje na temelju višekriterijskih procjena uključujući aspekte poljoprivrede, prinosa i kvalitete. Kao primjer, maksimalni sadržaj vlakana od 16,7%, kako je utvrđeno laboratorijskom opremom, može se postići za jedan od novih genotipova unutar prve godine testiranja na dva polja. Na kraju, ali ne i najmanje važno, spominje se istraživački projekt u kojem su od 2015. do 2018. istraženi aspekti uzgoja i poljoprivrede, kao što su aspekti prerade u lancu vrijednosti [26].
2.2. Prerada vlakana
Slika 2. prikazuje razvoj tehnologije ekstrakcije vlakana koprive od 18. stoljeća do danas. Prije tri stotine godina, standardna procesna linija za proizvodnju vlakana koprive za ručno predenje uključivala je sljedeće procesne korake: ručnu žetvu stabljika, sušenje na suncu, lomljenje ogoljenih stabljika, ispiranje i sušenje i konačno njihovo sjeckanje (linija; [27] cit. u [28] Druga ručna procesna linija za tekstil od koprive koja se i danas prakticira u zapadnom Nepalu je sljedeća ([29]; nije prikazano na slici 2): nakon ručne žetve biljke vlakna (Girardinia diversifolia—Urticaceae; “Allo fiber ” prema [30]) željeznim srpom otrljaju se listovi i oštre dlake, a kora se nakon ručnog lomljenja skida ručno neposredno na licu mjesta. Za odvajanje (degumiranje) kora prekrivena pepelom se kuha. u vodi četiri sata, nakon čega slijedi uzastopno udaranje lišća drvenim čekićem i ispiranje vodom. Nakon toga se snopovi vlakana pročišćavaju stavljanjem lišća u mješavinu gline i vode i, nakon sušenja na suncu, ponovnim udaranjem snopova obloženih glinom sa drvetom n čekić. Istegnuti i paralelizirani snopovi vlakana mogu se zatim koristiti za ručno predenje [29,31].
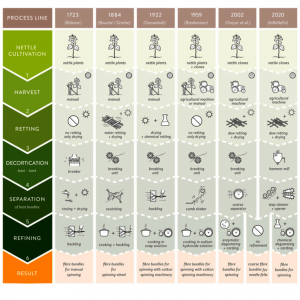
Slika 2 Uzdužni (Vanjska datoteka koja sadrži sliku, ilustraciju itd. Naziv objekta je materials-15-04288-i001.jpg) i neuređen (Vanjska datoteka koja sadrži sliku, ilustraciju itd. Naziv objekta je materials-15-04288 -i002.jpg) procesne linije za ekstrakciju i separaciju vlakana koprive od 1723. godine do danas.
Prije Prvog svjetskog rata, “tradicionalna” procesna linija za proizvodnju vlakana od koprive za predenje bila je vrlo slična proizvodnji vlakana od konoplje [32]. Nakon ručne žetve biljaka koprive, stabljike su se tradicionalno natapale u vodi ili u polju [9,32]. Močenje je proces koji koristi djelovanje mikroorganizama i vlage na biljke kako bi se otopio ili istrulio veći dio staničnih tkiva i pektina. Nakon sušenja i ljuštenja lišća, dekortikacija je obavljena pomoću jedinice za lomljenje, snopovi lična vlakna su odvojeni struganjem kako bi se nečistoće odvojile od sirovog materijala, a pročišćavanje je obavljeno kuhanjem i sjeckanjem (druga linija; [33] ). von Roeßler-Ladé preporučio je kuhanje vlakana koprive prije hakiranja [32]. To se razlikuje od prerade konoplje koja se kuhala tek nakon predenja. Ganswindt izvještava o povećanju proizvodnje vlakana koprive za njemačku tekstilnu industriju tijekom Prvog svjetskog rata (1914. – 1918.) jer su zalihe lana i pamuka bile potrošene [34]. Školarci su skupljali (“berli”) stabljike koprive i dobivali 6 do 14 D-Maraka (3-7 €) za 100 kg stabljika, ovisno o duljini [34]. Nakon sušenja i uklanjanja listova, izvršeno je kemijsko otapanje s otopinom amonijaka kako bi se razgradila pektinska lamela. Alternativno, provedeno je kvašenje vodom. Nakon toga, stabljike su ukrašene pomoću jedinice za lomljenje, a zatim su snopovi vlakana rascijepljeni, a vlakna su ispravljena. Nakon rezanja, izolirani, ali grubi snopovi vlakana koprive kuhani su u otopini sapuna kako bi se postigla kvaliteta vlakana koprive koja bi se mogla presti na strojevima za predenje pamuka. Tijekom nekoliko pokusa u svojoj predionici, direktor [35] je otkrio da su izolirana pojedinačna vlakna koprive sposobna za predenje. Ova procesna linija opisana je u [34] i prikazana je na slici 2 u trećem retku. Tijekom Drugog svjetskog rata (1939. – 1945.) ponovno su se povećala istraživanja korištenja vlakana koprive u njemačkoj tekstilnoj industriji. Bredemann je opisao dvije glavne metode koje su se razvile u to vrijeme: postupak jata bast (prikazan na slici 2, linija 4) i takozvani Elster postupak (nazvan po tekstilnom poduzetniku Johannesu Elsteru iz tvrtke Gebrüder Uebel u Njemačkoj) [16 ]. Stabljike koprive nisu bile namakane za postupak obrade, samo su osušene do određenog stanja (7-9% udjela vode), skinute im foliju i naknadno očišćena kora pomoću jedinice za lomljenje [36]. Nakon toga, ličje je očišćeno pomoću češljaste tresilice, kuhano u otopini natrijevog hidroksida (NaOH) i grebano prije predenja [16]. Nasuprot tome, Elster postupak uključuje različite procese kuhanja u vodi s odgovarajućim tvarima. Za cjelokupnu obradu korišten je stroj koji je zaštićen s nekoliko patenata [37,38]. Nažalost, ovaj stroj je tijekom posljednjih ratnih dana uništen [16].
Održivu alternativu za metode kemijske obrade predstavili su Dreyer i dr., koristeći proces degumiranja s enzimima za uklanjanje fosfatida iz sirovih ulja kako bi se poboljšala fizička stabilnost i olakšala daljnja obrada te postigla vlakna koprive s tekstilnom kvalitetom [39]. Dreyer i sur. [39] su u svojim istraživanjima koristili stabljike koprive natopljene u polju. Neuređena vlaknasta linija (ukupna vlaknasta linija) korištenjem čisto mehaničkog postupka (poznatog za konoplju; vidi [40]) korištena je za postupak dekortikacije i odvajanja, korištenjem jedinice za lomljenje i grubog separatora. Procesna linija koju su opisali Dreyer et al. [39] prikazano je na slici 2, linija 5. Enzimska obrada ima ekološke prednosti u usporedbi s kemijskom obradom [39]. Na temelju rada Dreyera i sur. [39], proizvodnja u mjerilu od otprilike 50 kg za enzimatsku obradu ličnatih vlakana provedena je poluindustrijski [41]. Industrijska procesna linija, koju danas koristi tvrtka NFC GmbH Nettle Fiber Company (Dahlenburg, Njemačka) za proizvodnju tekstilnih vlakana, opisana je u [26] i prikazana na slici 2, linija 6. Kultivirani klonovi koprive su strojno požnjeven, otopljen u polju i osušen. Za ukrašavanje se koristi mlin čekićar. Čišćenje je podijeljeno u dva koraka: tambur i čistač stepenica. Konačno, snopovi vlakana se pročišćavaju pomoću otvarača [26]. Za predenje fine pređe izvodi se degumiranje i grebenanje. Zanemarujući korake rafiniranja enzimatskog ili kemijskog degumiranja i grebanja (potrebno za predenje pojedinačnih vlakana koprive ili finih snopova vlakana u finu pređu; prikazano na slici 2, retke 5 i 6), mogu se proizvesti snopovi grubih vlakana za tehničke primjene poput iglanog filca. Po prvi put, mogućnosti poljoprivredne proizvodnje koprive s relativno velikim uzgojem i kvaziindustrijskom ekstrakcijom vlakana mogu se integrirati u okviru istraživačkog projekta [26]. Uspostavljeni (npr. klon B 13, “Z” klonovi), i po prvi put, novi “L” genotipovi su uzgojeni pod različitim gnojidbama i shemama gustoće sadnje kako bi se procijenili aspekti prinosa, obrade i kvalitete. Daljnji eksperiment o preradi vlakana pokazao je da vrijednosti sadržaja vlakana koje se nalaze u literaturi uvelike ovise o metodama ekstrakcije (npr. kemijski određen (“čisti”) sadržaj vlakana u odnosu na mehaničku obradu s laboratorijskim dekortifikatorima).
3. Biologija, fiziologija i genetika Urtica dioica L.
Cilj sljedećeg odjeljka je opisati trenutno znanje o biologiji, genetici i ekologiji koprive, s naglaskom na anatomiju stabljike te morfologiju i sastav ličnjaka. Ovo je znanje presudno za bolje razumijevanje različitih koraka u odabiru koprive, obradi slame i ekstrakciji vlakana, te za procjenu prinosa vlakana i analizu svojstava vlakana i prikladnosti za primjenu u tekstilu i materijalima.
3.1. Biologija, ekologija i reprodukcija
Urtica dioica nalazi se u mnogim hladnim do umjerenim regijama svijeta: Africi, Americi, Aziji, Australiji i Europi. Kopriva je rasprostranjena u sjevernoj Europi i Aziji, a manje u južnoj Europi i sjevernoj Africi [42]. Također je široko rasprostranjen u Sjevernoj Americi, posebno u Kanadi i Sjedinjenim Državama, a raste u izobilju na sjeverozapadu Tihog oceana, posebno tamo gdje je godišnja količina oborina velika. Prema Darwinovoj hipotezi prekooceanske difuzije, sjemenke koprive su održive nakon što su dugo plutale u morskoj vodi, pogodujući širenju na velike udaljenosti, što može objasniti njihovu široku geografsku rasprostranjenost [43]. Na razini podvrste ili sorte distribucija je vrlo različita. Na primjer, Urtica dioica subsp. gracilis izgleda da ima zemljopisnu rasprostranjenost ograničenu na Ameriku, dok Urtica dioica subsp. dioica je široko rasprostranjena [42]. Kopriva je zeljasta nitrofilna višegodišnja biljka koja raste na širokom spektru staništa, kao česta vrsta priobalnih staništa, močvara, livada, riječnih obala, pustara, poplavnih ravnica i narušenih područja. Zabilježene su velike monospecifične sastojine na rubnim mjestima (npr. gomile šljake, dvorišta) bogate hranjivim tvarima [44]. Također se često nalazi pod obalnim vrbama diljem Europe [45]. Preferira vlažna, bogata tla i može uspijevati na punom svjetlu, ali najbolje uspijeva u polusjeni. Prema Tayloru [12], Urtica dioica subsp. dioica ne može održavati anoksične uvjete dulje vrijeme, na primjer, kao što se može dogoditi s poplavama [46]. Kopriva preferira rahla tla s organskom tvari i visokom razinom dušika za brzi rast [47]. Dušik potiče rast nadzemnih dijelova, a smatra se da je dušik najvažnija komponenta ishrane koprive [48]. Vrsta je morfološki vrlo plastična [49] te stoga obuhvaća veliki broj podvrsta na svim kontinentima [42]. Međutim, sve podvrste su uspravne sa žućkastim i cilindričnim rizomima i stolonima (Slika 3a). Korijenov sustav se nalazi u organskom horizontu na malim dubinama (10 do 30 prvih cm). Stabljika ima četverokutni presjek (Slika 3b) i može doseći visinu do dva metra s listovima u nasuprotnim parovima (Slika 3c), ovalnim ili čak kopljastim, sa zaobljenom ili srčanom bazom, nazubljenim rubovima lista i šiljastim ili šiljastim listom vrhu (slika 3d). Dva nasuprotna lista i četiri stipule stabljike umetnute su u svaki čvor, osim za kotiledonarni čvor i prvi čvor glavne stabljike, koji nemaju stipule [12] (Slika 3e). Mladi listovi su organi koprive koji sadrže najviše vlage (dvostruko više nego u korijenu), zbog čega su posebno nježni [50]. Cvatovi su aksilarni, klasasti, po četiri u čvoru, s mnogo malih zelenih jednospolnih cvjetova [51]. Cvjetovi se pojavljuju od lipnja do listopada [47], s muškim i ženskim cvjetovima koji se obično nalaze u različitim biljkama (npr. Urtica dioica subsp. dioica). Međutim, prijavljene su dvodomne (Urtica dioica subsp. dioica) i jednodomne (Urtica dioica subsp. gracilis) podvrste [52]. Große-Veldmann i Weigend [53] također su predložili da nijedna striktna dvodomna vrsta ne pripada rodu Urtica. Urtica dioica i sve njezine podvrste su poligamne, pri čemu Urtica dioica subsp. dioica uglavnom zastupljena s 80 do 90% dvodomnih jedinki i 10% jednodomnih jedinki (bazalni muški cvatovi i vršni ženski cvatovi). Muški cvjetovi su uspravniji, žućkasti, s četiri duguljasta prašnika presavijena u cvjetni pupoljak, dok su ženski cvjetovi zelenkasti, s jednolokularnim jajnikom na vrhu s četkastim oblikom i žigom, te više vise [54,55] ( Slika 3f,g). Plodovi su ahenije, lagane (0,2 mg), vrlo sitne (1,3 × 1,0 mm2) i stoga ih lako nosi vjetar [56]. Drvenasta stabljika čini 23-30% ukupne biomase [57]. Međutim, čini se da abiotski čimbenici poput nadmorske visine uzrokuju morfološke i anatomske varijacije u populacijama koprive [58].

Slika 3 Morfologija koprive s fokusom na (a) korijenje, (b) dio stabljike, (c) izdanke, (d) list, (e) čvor, (f) ženske i (g) muške cvjetove, (h) list prekrivene oštrim dlakama i (i) oštrim dlakama na stabljici.
Kopriva je prekrivena dlačicama na listovima (Slika 3h) i na peteljkama (Slika 3i), koje se nazivaju trihomi, a predstavljaju ih kratke jednostavne dlake i duže krute dlake koje bodu. Gustoća trihoma manja je na dnu stabljike, u internodijama i na gornjoj površini lišća [59]. Kad puknu, male cjevčice ispuštaju tekućinu koja sadrži mravlju kiselinu (metanoičnu kiselinu; CH2O2), serotonin (5-hidroksitriptamin; C10H12N2O), histamin (2-(1H-imidazol-4-il)etanamin; C5H9N3) i acetilkolin ( 2-acetoksi-N,N,N-trimetiletanaminij; C7H16NO2) [60,61]. Ovi spojevi uzrokuju svrbež i peckanje te tako služe kao obrambeni mehanizam protiv insekata, sisavaca biljojeda ili velikih paša [5]. Taj je mehanizam i mehanički i biokemijski [62]. Postoje dokazi da su trihomi evoluirali za obranu koprive od sisavaca biljojeda [63]. Doista, populacije pod intenzivnom ispašom sisavaca imaju više trihoma nego populacije u područjima s manje intenzivnom ispašom [64]. Fenotipska plastičnost je važna osobina za Urtica dioica; na primjer, proizvedeno je manje trihoma kada su rasli u sjeni nego na suncu u eksperimentu kulture [65]. Ovi eksperimenti su također postavili hipotezu da su te varijacije (uključujući polimorfizam u gustoći kose) genetske i nasljedne [66]. Glavna strategija razvoja koprive je podzemno vegetativno razmnožavanje. U kasno ljeto, kada dođe do opadanja listova, stabljike se ulegnu i tvore rizom. Nova jedinka može se proizvesti iz svakog čvora stabljike ili starijih rizoma [44]. Izbojci iz rizoma razvijaju se uglavnom u jesen, tijekom zime, i nastavljaju s rastom sljedećeg proljeća [67], iako neki mogu umrijeti. Otprilike jedna trećina maksimalne biomase izdanaka održava se tijekom zime [44]. Dreyer [17] je iz svog istraživanja zaključio o izrazitom ranijem rastu izdanaka u kasnu zimu/proljeće na temelju relativno velikog broja izdanaka kao i velike količine hranjivih tvari u rizomu. Ova reproduktivna strategija objašnjava invazivni karakter koprive, formirajući guste monospecifične ili čak monoklonske sastojine u okruženjima gdje druge biljke imaju nisku kompetitivnu sposobnost ili kada su uvjeti tla povoljni. Tako su mjerenja duljine korijena pokazala da se kopriva može razviti više od trećine više u odnosu na npr. ječam, zob ili grah u gornjem sloju od 25 cm razmatranog sloja [17]. Kopriva se može razmnožavati i spolnim putem. Međutim, spolno razmnožavanje ima mali utjecaj na njegovo širenje, ali je opisano kao bitno za kolonizaciju novih mjesta [68]. Jedna biljka koprive može proizvesti do 20 000 sjemenki na otvorenim površinama, dok se taj broj smanjuje na 5000 u zasjenjenim područjima [69]. Sjeme se obično sije od kolovoza [70], a sjeme koprive može ostati održivo dugo vremena u slanoj vodi (do 240 dana) [43] i preživjeti gutanje od strane životinja [12]. Širenje sjemena koprive ne temelji se na jednoj prevladavajućoj strategiji, već na nekoliko mehanizama (npr. vjetar, voda, kukci) koji su više ili manje učinkoviti ovisno o okolišu biljke [12,70]. U umjerenim klimatskim uvjetima klijanje obično počinje početkom siječnja i doseže vrhunac u travnju [12]. Klijanje je inhibirano mrakom, a stimulirano svjetlom i temperaturnim fluktuacijama [71], tako da spolno razmnožavanje nije vrlo učinkovito na mjestima s visokim vegetacijskim pokrovom.
3.2. Morfologija stabljike i vlakana i sastav vlakana
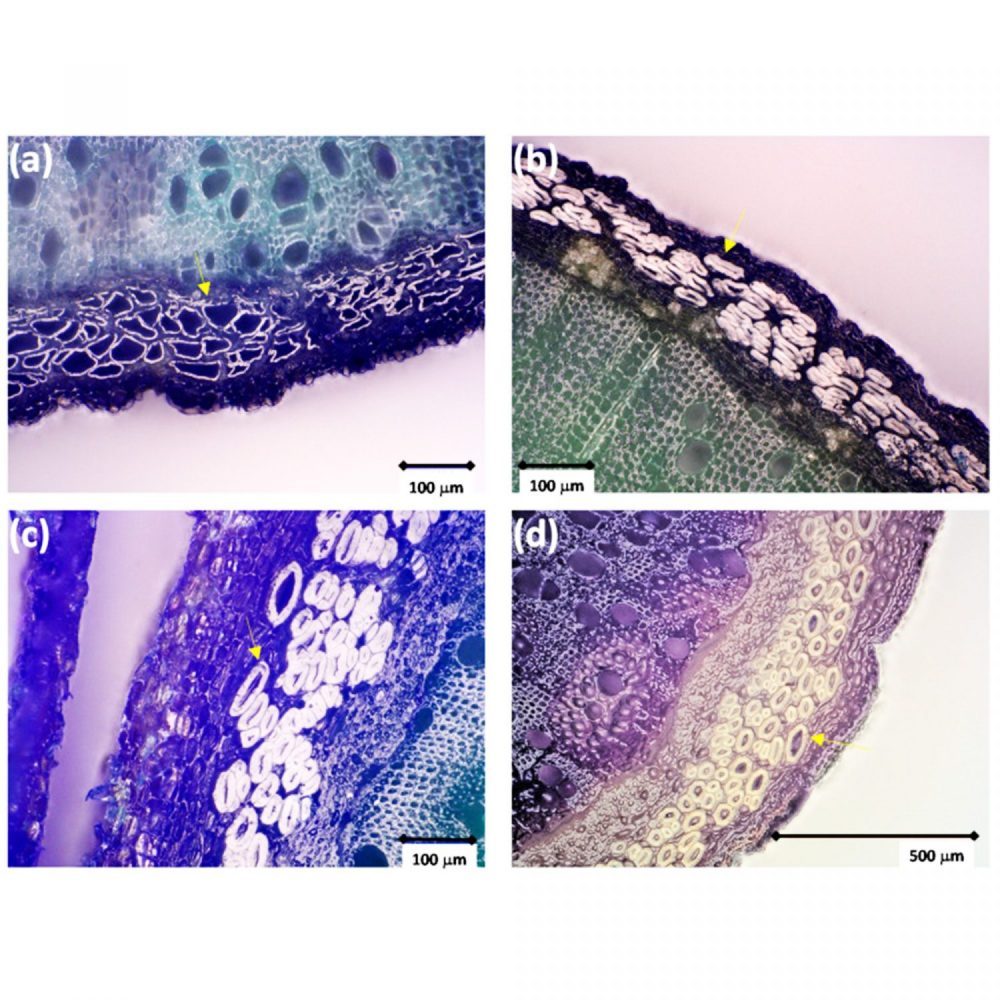
U otvorenoj literaturi, najdetaljniji opis anatomije stabljike koprive i, šire, vegetativnih organa koprive nedvojbeno je u memoarima Augustea Gravisa objavljenim 1885. [72] kao što je prikazano u dodatnom materijalu na slici S1. Temelji se na ispitivanju velikog broja stabljika, lišća i korijena, uključujući ne manje od 15 000 rezova (poprečnih, ali i radijalnih i tangencijalnih) napravljenih na različitim stupnjevima razvoja, u različitoj dobi i uzimajući u obzir uvjete rasta. Ako je ovo istraživanje izvorno trebalo poslužiti kao osnova za botaničku klasifikaciju, ono danas predstavlja jednu od najdetaljnijih studija o anatomiji koprive. Posljednjih su godina lična vlakna bila predmet više karakterizacije zbog njihove potencijalne primjene u tekstilu ili drugim materijalima. Primjerice, analize su napravljene na divljoj koprivi u okviru projekta PHYTOFIBER (www.phytofiber.fr (pristupljeno 1. rujna 2018.), [11]). Slika 4 prikazuje nekoliko primjera tipičnih poprečnih presjeka stabljika. Promjer primarnih ličnih vlakana kretao se od nekoliko mikrona do najviše 100 µm, s debljinom stijenke od jednog µm do više od 20 µm, ovisno o starosti, položaju stabljike i zrelosti biljke. U skladu s opažanjima Gravisa [72], može se vidjeti (Slika 4b) da su primarna lična vlakna ponekad kolabirana, ovisno o debljini stanične stijenke kada dijametralni rast počinje u stabljici. U analiziranim stabljikama nisu uočena sekundarna lična vlakna. Bacci i sur. [2] izvijestio je za lična vlakna kultivirane koprive (njemački klon vlaknaste koprive 13), srednje promjere od 19, 32 i 47 µm u gornjem, srednjem i donjem dijelu stabljike s prosječnom duljinom od 58, 50 µm. , i 43 mm u tim istim dijelovima. Što se tiče njihovog biokemijskog sastava, Dreyer i Edom [73] vrijednosti prikupljene iz literature s približno 54% celuloze, 10% hemiceluloze (uglavnom sastavljene od arabinana, ksilana, galakturonana), 4,1% pektina, 9,4% lignina, 4,2% vosak i masti, te 18% proizvoda topivih u vodi. Na ovaj kemijski sastav uvelike utječe kvašenje ili različite metode koje se koriste za ekstrakciju vlakana [9,74] i datum berbe [2]. Nakon kvašenja udio celuloze može doseći vrijednosti do 88%, dok udio ostalih sastojaka pada na 4%, 0,6%, 5,4%, 3,1% odnosno 2,1%.
Slika 4 Poprečni presjeci stabljika divlje koprive ubrane u Saint-Symphorienu (Francuska, regija Bourgogne-Franche-Comté (lat. 47°5′5.98″ N. 5°19′44.0322″ E) 2019. u različito vrijeme između svibnja (a ), lipanj (b), srpanj (c) i kolovoz (d). Strelice pokazuju neke primjere primarnih ličnih vlakana (a–c): Placet et al., neobjavljeni podaci; (d), [11]).
3.3. Filogenija i genetske značajke
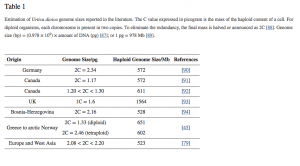
Rod Urtica je vrlo karakterističan i lako ga je identificirati, ali je razgraničenje vrsta još uvijek problematično, posebno za Urtica dioica L., koja ima više od 20 infraspecifičnih svojti priznatih u Euroaziji i Americi [75,76]. Große-Veldmann i Weigend [77] identificirali su pet morfotipova Urtica dioica subsp. dioica: var. dioica, var. hispida, var. sarmatica, var. Holosericea, i var. glabrata. Ovi se morfotipovi razlikuju u odnosu na preferencije staništa, geografsku rasprostranjenost u Europi, varijacije indumentuma (npr. gustoća trihoma i broj dlačica koje bode), oblik lista i morfologiju ruba lista (npr. blago ili većim dijelom ovalna, srcasta ili odrezana baza) [42,78]. Razina ploidnosti je drugi razlog koji opravdava prepoznavanje infraspecifičnih svojti. Poliploidija oblikuje obrazac ograničavajući diploidne citotipove na rezidualna staništa [79]. U dvije najnovije filogenije [42,80], Urtica dioica s.l. tvori dobro poduprtu klasu koja se sastoji od različitih povezanih taksona, bez obzira na korištene markere (nuklearne, kloroplastične ili kombinaciju ova dva tipa). Ovaj klas spada u zapadni euroazijski klas i azijsko-američki klas. Točnije, zapadna euroazijska skupina uključuje sve U. dioica s.str. i pokazuje sestrinsku vezu s endemičnom mediteranskom skupinom (tj. U. atrovirens, U. bianorii i dioica ssp. Cypria) i dvije srodne afričke U. massaica i U. simensis. Drugi klas grupira zapadno- i sjevernoamerički U. gracilis u subkladu sestru azijsko-australskog subklada. Međutim, različite podvrste, ili čak varijeteti, predstavljaju morfološke razlike koje bi mogle biti posljedica fenotipske plastičnosti, a ne genetske divergencije [80]. Rejlová i sur. [79] stoga sugeriraju da veličina genoma može doprinijeti razgraničenju i otkrivanju blisko povezanih vrsta (npr. Urtica bianorii i Urtica kioviensis pokazale su veće veličine genoma u svojoj studiji). Razlike u vrijednostima veličine genoma mogu ukazivati na genetsku udaljenost. Međutim, veličina genoma ne omeđuje podvrstu, već je samo razina ploidnosti prihvaćena kao razgraničavajuća značajka Urtica dioica subsp. dioica. Filogenetski podaci dobiveni korištenjem molekularnih markera pokazuju da morfološke i geografske karakteristike koje se koriste za razlikovanje i grupiranje vrsta ne odražavaju filogenetsku povezanost [81]. Standardni molekularni markeri ne rješavaju odnose na razini podvrste ili sorte. Große-Veldmann [80] je stoga upotrijebio pristup genotipizacije po sekvenciranju (GBS) na 53 taksona. Koristili su rezni enzim PstI-HF (mjesto prepoznavanja: CTGCA’G), a enzim MspI osjetljiv na metilaciju (mjesto prepoznavanja: C’CGG) također je korišten za razumijevanje evolucijskih odnosa unutar kompleksa vrsta koje pripadaju rodu Cycnoches, a tropske orhideje [82], te zaključiti filogeniju sedam blisko povezanih vrsta iz roda Carex [83]. Dobili su 4013 lokusa i 30.840 SNP-ova. Međutim, to nije omogućilo identifikaciju infraspecifičnih odnosa Urtica dioica sensus stricto. Odvojeni tretman različitih alela također nije poboljšao rezoluciju. Prethodni rezultati studija filogenetskih odnosa na temelju standardnih nuklearnih markera (npr. ITS, trnS-trnG, trnL-trnF, psbA-trnH) dali su uglavnom iste rezultate. Farag i sur. [84] pokazali su da postoji mala sličnost između 43 skupine sekundarnih metabolita (uglavnom fenolnih spojeva i hidroksilnih masnih kiselina) i filogenetskih podataka. Međutim, jedna podskupina pronađena je u obje analize: Urtica dioica, koja se pojavljuje kao isključiva skupina. Većina objavljenih procjena veličine biljnog genoma napravljena je protočnom citometrijom [85]. Ova se metoda sastoji od procjene sadržaja DNA izoliranih jezgri obojenih DNA-selektivnim fluorokromom [86]. Sadržaj DNA u haploidnoj stanici obično se mjeri C-vrijednošću izraženom u pikogramu (pg), pri čemu 1 pg iznosi oko 978 Mb [87]. Za Urtica dioica s. str., procjene veličine genoma kreću se od 597 do 1540 Mbp (Tablica 1) [80].
3.4. Fitokemija koprive
Različiti metabolomski pristupi (npr. plinska kromatografija-masena spektrometrija (GC-MS), ultra-high-performance tekuća kromatografija-visoka rezolucija masena spektrometrija (UHPLC-HRMS/MS)) primijenjeni su za otkrivanje sadržaja metabolita u Urtica dioica . Razni spojevi koji mogu imati prehrambenu i/ili medicinsku važnost otkriveni su ovim pristupima probira. Shokrzadeh i sur. [95] pokazali su da ekstrakti Urtica dioica imaju terapeutski potencijal za ublažavanje oksidativnog stresa i hiperglikemije izazvane dijabetesom. Veliki broj spojeva mogao bi biti odgovoran za te učinke. Doista, veliku raznolikost sekundarnih metabolita otkrili su Al-Tameme i sur. [96], koji su prijavili aromatske prstenove, alkene, alifatski fluor, alkohole, etere, karboksilne kiseline, estere, nitro spojeve, alkohole s vodikovom vezom i fenole u metanolnim ekstraktima Urtica dioica. Isto tako, alkaloidi, saponini, tanini, flavonoidi, steroidi i terpenoidi, polifenoli i srčani glikozidi otkriveni su u lišću Urtica dioica na temelju infracrvene (FTIR) spektroskopije s Fourierovom transformacijom [97]. Pinelli i sur. [98] izvijestili su da klorogenska i 2-O-kafeoil jabučna kiselina dominiraju fenolnim spojevima u lišću, dok su u peteljkama pronađeni uglavnom flavonoidi i antocijanini. Točnije, Grauso et al. [99] otkrili su da ekstrakti koprive sadrže dva pentaciklička triterpenola α- i β-amirin u nepolarnoj frakciji, dok su u polarnom ekstraktu pronađene velike količine kolina. Brahmi-Chendouh i sur. [100] izvijestili su da su derivati hidroksicimetne kiseline uz C-glikozilirane flavone najreprezentativniji sastojci lišća koprive. Međutim, čini se da svojte koje pripadaju Urtica dioica imaju sličan sastav metabolita i, prema tome, slična farmakološka svojstva [84]. Suvremene hidroponske metode uzgoja u staklenicima također su postavljene za lakše upravljanje okolišnim čimbenicima kako bi se poboljšala proizvodnja metabolita [101]. Ipak, mužjaci i ženke imaju različit sadržaj i kemijski sastav polifenolnih kiselina u lišću, pri čemu je muški oblik karakteriziran većim sadržajem ovih spojeva [102]. Na fenolne spojeve u lišću koprive snažno utječu stanište i nekoliko drugih biotičkih i abiotskih čimbenika [103,104]. Ukupni sadržaj fenola također je različit ovisno o fenološkom stadiju koprive [103,105]. Čini se da se najveći sadržaj polifenola javlja između travnja i srpnja na početku vegetacijskog razdoblja [106,107]. Točnije, smanjio se u lišću od proljeća do jeseni, dok je blagi porast primijećen u korijenu [108]. Nedavni rad istaknuo je činjenicu da su biološki resursi samoniklih vrsta Urtica dioica L. s europskog juga Rusije vrijedan izvorni materijal za dobivanje sorti s vrijednim biokemijskim svojstvima [109]. Kao biljka bogata silicijem, kopriva također predstavlja vrijedan interes za kozmetiku i prirodnu medicinu [110]. Naprotiv, drugi spojevi kao što su alergijski proteini su pronađeni i mogu uzrokovati rinitis kod ljudi kao što je prikazano korištenjem alergomičkog pristupa; međutim, potrebna su daljnja istraživanja kako bi se procijenio alergenski potencijal Urtice dioice [111].
>Literaturna izvješća o koncentracijama Ca, Mg i K u biomasi u Urtica dioica variraju (tablica S1 dopunskog materijala). U [112], K je bio glavni skupni element u lišću (33,9 g kg-1), a slijede ga Ca (28,6 g kg-1) i Mg (8,69 g kg-1), unutar istog reda veličine kao one objavljene u nedavnom radu [113]. U Kari [114], Ca je bio glavni makronutrijent u biljnoj infuziji koprive (naizgled lišće, 38,4 g Ca kg−1, 17,5 g K kg−1, 7,32 g Mg kg−1), dok je [115] izvijestio veće koncentracije Mg u lišću Urtica dioica, u rasponu od 25,1-35,6 g kg−1. Uspoređujući elementarni sastav stabljike i lišća, Mg i Ca rezultirali su 2 odnosno 3 puta većom zastupljenošću u lišću nego u stabljikama Urtica dioica [115]. Za željezo se navodi da je najvažniji TE (element u tragovima) u koprivi [112] i glavni TE u lišću, s vrijednostima u rasponu od 151 mg kg-1 [112] do 999 mg kg-1 [114]. U usporedbi s koncentracijama TE u lišću biljaka [116], koncentracije Cu, Zn, Cr i Co u koprivi za lišće i cijele biljke [11,112,114,115] mogu se smatrati unutar fizioloških razina kada se uzgajaju u nezagađenim tlima; iznimke su zabilježene u [117], gdje je koncentracija Zn i Cr u cijelim biljkama Urtica dioica premašila fiziološke vrijednosti u lisnatim usjevima [116]. Koncentracije mangana u koprivi značajno variraju ovisno o različitim mjestima uzorkovanja. Za uzorke iz Makedonije utvrđen je manjak Mn na svim analiziranim lokacijama [115] u usporedbi s fiziološkim koncentracijama Mn u biljkama [116], dok su u uzorcima iz Belgije [117] i Srbije [112] koncentracije Mn u biomasi koprive bile u fiziološki raspon. Pb, As i Hg u Urtica dioica znatno su ispod razina “toksičnih” biljaka [116] u svim literaturnim vrijednostima, s iznimkom napravljenom za Pb u uzorcima koprive iz nekontaminiranog tla u Belgiji (34 mg kg−1, [117] ).
3.5. Organizmi povezani s koprivom
Dostupno je malo informacija o organizmima povezanim s korijenom koprive [118]. Toubal i sur. [119] proučavali su bakterijsku raznolikost povezanu s različitim tkivima koprive pomoću biokemijskih testova i spektrometrijskih analiza (MALDI-TOF MS), što je rezultiralo izolacijom 7 rodova i 11 vrsta koje pripadaju rodovima Bacillus, Escherichia, Pantoea, Enterobacter, Staphylococcus, Enterococcus i Paenibacillus. Za različita tkiva koprive, najčešća identificirana vrsta je Bacillus pumilus. Također se pokazalo da je kopriva domaćin Ralstonia solanacearum, koja uzrokuje smeđu trulež krumpira [120]. Mojičević i sur. [121] izolirani Streptomyces spp. iz rizosfernog tla U. dioica, koji ima sposobnost stvaranja antifungalnih spojeva protiv Candide krusei, C. parapsilosis i C. glabrata. Mikroskopska promatranja obojenih (Trypan blue) i obilježenih (WGA-AF488) segmenata korijena U. dioica otkrila su da su gljivične strukture kolonizirale kortikalne stanice, što uključuje mikrosklerocije formirane od tamnih septiranih endofita i spora (Slika 5). Druga opažanja otkrila su relativno nisku stopu AMF struktura u korijenju [122]. Djelomična raznolikost gljivičnog mikrobioma koprive koja je povezana s korijenom uzgojene na odlagalištu sedimenta obogaćenog metalima Fresnes-sur-Escaut nedavno je karakterizirana korištenjem tehnološkog pristupa Illumina MiSeq [123]. U mikobiomu koprive dominiraju Pezizomycetes i Leotiomycetes, uključujući endofitne i saprotrofne taksone (Slika 5), pri čemu je navodno saprotrofni rod Kotlabaea bio najzastupljeniji. Po raznolikosti i brojnosti najzastupljeniji su redovi Pezizales, Helotiales, Pleosporales, Agaricales, Hypocreales i Thelephorales [123]. Samo 54% sekvenci gljivica uspješno je pripisano rodu, što odražava nedostatak podataka o mikrobiomu koprive. Uz Kotlabaea, Olpidium, Tetracladium i Hymenoscyphus bili su među najobilnijim identificiranim rodovima. Unatoč tome što je poznata biljka koja nije AMF, kopriva je povezana sa značajnim udjelom ektomikoriznih OTU (9,7%), što ukazuje na neke veze sa simbiotičkim mikobiomom okolnih topola [123].
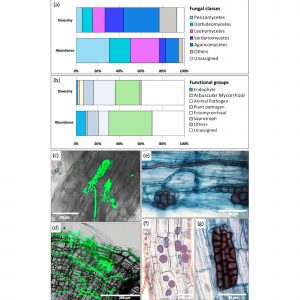
Slika 5 Relativna raznolikost i brojnost (%) najzastupljenijih razreda gljiva (a) i funkcionalnih skupina gljiva (b) povezanih s korijenjem koprive s lokaliteta Fresnes-sur-Escaut. Klase s relativnom zastupljenošću <5% i manje zastupljene i raznolike funkcionalne skupine okupljene su u skupini "ostalo" (prilagođeno iz [123]). Strukture endofitnih gljiva promatrane fluorescentnom (c,d) i fotonskom (e–g) mikroskopijom u pripravcima korijena koprive označenog s WGA-AF488 ili obojenog tripan plavim. (c) gljivične hife koje koloniziraju kortikalnu stanicu; (d) mreža izvanstaničnih hifa gljiva; (e) hife koje tvore mikrosklerocije poput mozga; (f) unutarstanične spore gljivica različite morfologije; (g) melanizirani potpuno zbijeni mikrosklerocij (Yung et al., neobjavljeni podaci).
Prvi rad na kukcima koprivnjaču obavljen je u Europi 1970-ih [124,125,126] i istaknuo je njezinu važnost kao rezervoara insekata, ugošćujući veliku raznolikost, a posebno neke vrste Hemiptera i Coleoptera [127]. Kopriva je domaćin više od 100 vrsta insekata, od kojih se tridesetak klasificiraju kao specijalisti, uključujući nekoliko vrsta Aphididae, Psyllidae i Nymphalidae. Pronađene su mnoge druge generalističke vrste koje pripadaju obiteljima Miridae, Lygaeidae ili Cicadellidae, kao i kompletan skup grabežljivaca i parazitoida kao što su vrste Coccinellidae, Syrphidae i Braconidae. U novije vrijeme, studije provedene u Sjedinjenim Državama [128] i Belgiji [129,130,131] potvrdile su da je održavanje U. dioica kao mrlja ili učinkovitije kao monospecifične sastojine [132] pridonijelo poboljšanju bioraznolikosti unutar agrosustava. Kopriva bi posebno mogla promovirati korisne kukce, uključujući nekoliko taksona grabežljivaca (npr. Coccinellidae, Syrphidae, Anthocoridae) i parazitoida (npr. Braconidae, Diapriidae), poznatih kao prirodni konkurenti štetnicima usjeva [129,131,133]. Promicanje prirodnih neprijatelja potencijalnih vrsta štetnika povezano je s ugošćavanjem Microlophium carnosum, lisne uši koja služi kao odvraćanje ili alternativni plijen za mnoge predatore prisutne na obližnjim zaraženim usjevima [134]. Osim toga, pokazalo se da plantaža klona koprive odabranog zbog sadržaja vlakana može poduprijeti ovu vrstu lisnih uši i tako promovirati njezine prirodne neprijatelje [135]. Lisne uši čiji je domaćin kopriva mogu biti izvor hrane koji može privući lisne uši ili parazitoidne kukce [133]. Kako bi potaknuli migraciju predatora povezanih s koprivom u okolne agrosustave, Alhmedi et al. [129] predložili su žetvu koprive brzo nakon dolaska lisnih uši. Kopriva bi stoga mogla neizravno pridonijeti regulaciji štetnika usjeva u okolnom okolišu [131]. Sukcesija je proces motiviran pozitivnim interakcijama između vrsta, s pionirskim vrstama koje olakšavaju kolonizaciju okoliša vrstama koje su manje tolerantne na stres, ali su konkurentnije [136]. Stoga facilitacija predstavlja najvažniji proces u sukcesijskim sekvencama, posebno u primarnoj sukcesiji, gdje su okolišni uvjeti teški [137]. Tijekom primarne sukcesije, dušik u tlu je često ograničavajući faktor u uspostavljanju vegetacije [138]. Urtica dioica opisana je u literaturi kao vrlo konkurentna vrsta kada su uvjeti vlage i tla povoljni, posebno zahvaljujući svojoj sposobnosti da se vegetativno učinkovito razmnožava stolonima i rizomima. U usporedbi s drugim biljkama (npr. Agropyron repens, Artemisia vulgaris, Calamagrostis epigeios, Cirsium arvense, Epilobium angustifolium, Phalaris arundinaceae, Typha spp.), kopriva je pokazala najveću sposobnost širenja [139]. Međutim, kada uvjeti tla nisu optimalni, mnoge biljke kao što su čičak, čičak, Taraxacum officinale [140] ili Gallium aparine [141] mogu inhibirati njegov razvoj. Osim toga, studija o uzgoju Urtica dioica L. s drugim biljnim vrstama istaknula je da se kopriva dobro udomaćila u tlu koje je izvorno bilo zasijano mahunarkama, sa značajnim udjelom divlje bijele djeteline. Međutim, njegova sposobnost da se udomaći znatno je smanjena u okolišima u kojima dominiraju neke trave [142]. To ukazuje da je obrada tla prije sadnje važan čimbenik u razvoju ove biljke, iako se Urtica dioica smatra korovom u intenzivnoj poljoprivredi [9]. Nekoliko studija nastojalo je dokazati alelopatski potencijal koprive, koji ovisi o korištenoj koncentraciji i biljnoj vrsti. Na primjer, neke su studije pokazale inhibicijski učinak vodenih ekstrakata koprive na klijanje ječma [143], vjetrovke i janjeće četvrti [144], dok je [145] pokazalo da ekstrakti lista i korijena Urtica dioica značajno smanjuju masu korijena. Khan i sur. [146] otkrili su snažan inhibitorni učinak metanolnih ekstrakata koprive na klijanje i rast rotkvice. Nedavne genetske studije pokazuju da Urtica dioica prolazi kroz veliki protok gena što rezultira visokom stopom rekombinacije i da se morfološke razlike mogu pratiti unatrag do lokalne usmjerene selekcije i fenotipske plastičnosti. Molekularni pristupi koji se temelje na metabarkodiranju okoliša dalje omogućuju dešifriranje gljivičnog mikrobioma koprive, kojim su dominirali Pezizomycetes i Leotiomycetes. Ovaj se pristup proširio na velik broj različitih mjesta kako bi se bolje opisao temeljni mikrobiom koprive.
4.3. Vađenje i obrada vlakana
Iako su metode ekstrakcije korištene u prošlosti bile vrlo različite, opći postupak ekstrakcije vlakana koprive (ličje) može se dalje podijeliti u sljedećih pet procesnih koraka: žetva, otapanje, dekortikacija, odvajanje i rafiniranje (Slika 8). Ovaj odjeljak predstavlja pregled mogućih metoda u svakom koraku procesa na slici 8.
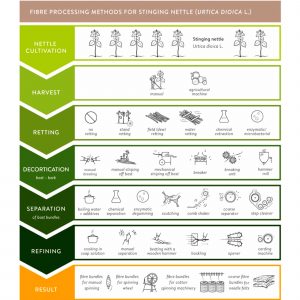
Slika 8 Metode ekstrakcije i separacije libnih vlakana za biljke koprive (kompilacija tehnika koje grafički ilustriraju metode opisane u tekstu).
5.4. Korištenje koprive u strategijama fitomenadžmenta
Potencijalno vrijedna sinergija za korištenje koprive za vlakna je to što se kopriva može uzgajati kao dio strategije upravljanja zemljištem za marginalno ili braonfild zemljište. Fitoupravljanje je pristup koji koristi biljke za stabilizaciju ili izvoz kontaminanata iz tla uz ograničavanje širenja i rizika od kontaminanata iz tla [219]. U fitomenadžmentu se može koristiti nekoliko procesa, kao što su fitoekstrakcija, fitostabilizacija, fitovolatilizacija ili rizofiltracija [220]. U mnogim laboratorijskim eksperimentima u posudama, koncentracije mnogih TE u listovima koprive uzgojene na kontaminiranom tlu premašuju razine toksičnosti koje je prijavio Kabata-Pendias [116] i u Dodatnoj tablici materijala S3. To je slučaj, na primjer, za krom (Cr) [221, 222], selen (Se) [223], fluor (F) [224], arsen (As) [225], cink [221], olovo (Pb) , nikal (Ni) i kadmij (Cd) [226]. Ova sposobnost nakupljanja TE predložena je kao učinkovit adsorbent za uklanjanje npr. Cu2+ iz vodenih otopina nakon nepotpunog spaljivanja. Stoga bi to bio koristan potencijal biosorbensa za uklanjanje TE (npr. kadmija) iz otpadnih voda [227,228]. Ovaj kapacitet tek treba dokazati za in situ slučajeve. Doista, u lišću koprive prikupljenom in situ na kontaminiranim mjestima, razine Cd, Cu, Hg, Ni, Pb, Zn [11,229,230,231,232,233] i Mn [233,234] bile su znatno ispod toksičnih razina koje je prijavio Kabata-Pendias [116] . Međutim, razine TE u korijenu koprive obično su puno više od onih izmjerenih u lišću (Tablica dodatnog materijala S3). Očito, zbog potencijalnih koncentracija metala, kopriva uzgojena na zagađenim zemljištima nije prikladna za medicinske svrhe ili konzumaciju [235]. Uključivanje koprive u strategije upravljanja biljnim biljkama također donosi značajne prednosti bioraznolikosti. Nedavni pristupi optimiziranju sustava fitomenadžmenta ističu važnost sklopova između usjeva i spontanih vrsta [219] kako bi se pogodovala ekološka rehabilitacija ovih rubnih zemljišta [236]. Urtica dioica doista se često opaža kako spontano raste pod vrstama salicaceous [45], uključujući plantaže postavljene u kontekstu fitoupravljanja [43]. Taksonomska i funkcionalna raznolikost insekata povezanih s koprivom bila je usporediva s onima u prirodnom okolišu, naglašavajući da kopriva također djeluje kao rezervoar insekata u slučaju onečišćenog mjesta. Kao dio projekta fitomenadžmenta povezanog s proizvodnjom vlakana koprive [237], okarakterizirana je entomofauna povezana s agrosustavom kopriva-topola koji se nalazi na odlagalištu zagađenom Hg, te je određena izloženost insekata Hg. Kada se uzmu u obzir životne karakteristike kukaca, kukci vezani uz koprivu primarno su bili izloženi Hg kroz hranidbenu mrežu sa značajnim biomagnificiranjem, osobito na razini sekundarnih predatora. Doista, unutar hranidbene mreže povezane s koprivom, ukupne koncentracije Hg porasle su na sljedeći način: koprive < biljojedi < stručnjaci za grabežljivce < generalni grabežljivci, s jazom između stručnjaka za grabežljivce (uključujući Coccinellidae) i generalnih grabežljivaca, što ukazuje na vjerojatan unos Hg iz vanjskih izvora biovektori na ovoj razini [238]. Za razliku od insekata povezanih s koprivom, čini se da kontaminacija ne utječe na gljivičnu biomasu koprive, ali struktura zajednice može biti [118]. Međutim, u kontaminiranom okolišu dodavanje ostataka koprive (nadzemni dijelovi) dovodi do značajnog povećanja mikrobne biomase, sadržaja C, P, N i ergosterola, a time i biomase gljiva u tlu [118]. Uključivanje koprive u prakse upravljanja biljnim vrstama inovativno nudi potencijalne dobitke u pogledu održivosti, uključujući (i) obnavljanje prirodnih vlakana, (ii) samoodrživost/održavanje pokrova tla, (iii) njezinu visoku vrijednost bioraznolikosti kao izvora za raznolike prehrambene mreže, (iv) njezinu vrijednost odvraćanja od oštećenja posjetitelja na sustavima fitoupravljanja (zadržava ljude na stazama) i (v) njegovu toleranciju na širok raspon uvjeta tla, kao što su tla kontaminirana TE.
6. Zaključci
Kopriva je jedan od najstarijih izvora biljnih vlakana koje je čovječanstvo koristilo, ali je uglavnom izblijedjela iz komercijalne upotrebe u 20. stoljeću, osim kao ratni materijal. Međutim, kopriva ima mnoga svojstva koja dovode do ponovnog porasta interesa za korištenje vlakana koprive u 21. stoljeću. Koprivu je relativno lako uzgajati, osobito u umjerenim klimatskim područjima, sve dok ima dovoljno kiše. Oni su kozmopolitska vrsta koja se prirodno pojavljuje u mnogim regijama diljem svijeta i stoga podržavaju vrlo širok raspon ekoloških niša, posebno za beskralješnjake. Također su relativno otporne na štetočine i bolesti. One su trajnice, bitno smanjuju potrebu za sjetvom i izbjegavaju obradu tla. Štoviše, mogu se uzgajati na marginalnom zemljištu, uključujući područja zagađena organskim ili anorganskim zagađivačima, i mogu potencijalno igrati ulogu u smanjenju rizika od onečišćenja za ljude, vodu i ekološke receptore. Primjena koprive može biti sinergijska sa šumarstvom, primjerice proizvodnjom topole i koprive. Ovo je također vrlo zanimljivo za sanaciju jer se topole naširoko koriste u sustavima fitoremedijacije. Osim toga, kopriva je jestiva i sadrži širok raspon sastojaka i nutritivnih spojeva. Funkcionalnost uzgoja koprive, dakle, uključuje kombinacije sljedećeg: Oporaba prirodnih vlakana za upotrebu u kompozitnim materijalima i tehničkom tekstilu Oporavak sastojaka i nutraceutika Zajednički uzgoj s drvećem Upravljanje rizikom za mjesta zahvaćena onečišćenjem tla Povrat korištenja rubnog zemljišta Poboljšanje tla i sekvestracija ugljika u tlu Pružanje širokog spektra ekoloških niša za mnoge autohtone vrste (podupirajući šire pružanje usluga ekosustava). Iskorištavanje ovih funkcija je u povojima i presudno ovisi o dobrom razumijevanju temeljne biologije koprive, njezine strukture i uzgoja. Ovaj pregled detaljno uspoređuje trenutno stanje znanja o koprivi i njenom uzgoju kao osnovnom resursu za daljnji razvoj izvanrednih mogućnosti koje nam kopriva nudi kao resurs i za održivo upravljanje i poboljšanje zemljišta. Velik dio ovih informacija u ovom radu prilično je star i do sada nije bio uspoređen na način koji bi omogućio korištenje široj (online) istraživačkoj zajednici. Gledajući naprijed, postoji nekoliko područja u kojima bi se znanje i tehnike uzgoja i upotrebe koprive mogli dodatno poboljšati kako bi se omogućile brojne mogućnosti koje ovaj usjev pruža. To uključuje: Veći raspon podataka o sastavu vlakana koprive i testiranja funkcionalnosti za suvremene primjene (na primjer, u ojačanju za kompozitne materijale) Bolje razumijevanje varijabilnosti ovih vlakana i kako ta varijabilnost utječe na njihovu upotrebu Bolje razumijevanje utjecaja klime na rast koprive i svojstva vlakana koprive Dublje razumijevanje prehrambenih proizvoda i proizvoda od sastojaka dostupnih iz koprive i potencijal za isporuku tih proizvoda paralelno s prirodnim vlaknima iz iste požnjevene biomase Veće funkcionalno razumijevanje ekoloških posljedica proizvodnje i uporabe koprive Veći napor u pilotiranju i demonstraciji proizvodnje i uporabe koprive, a posebno na rubnim područjima, uključujući ona zahvaćena onečišćenjem tla Snažniji temelj za razumijevanje ekonomskih i širih posljedica održivosti proizvodnje i upotrebe koprive te potencijalnog doprinosa koji bi to moglo imati za rješavanje trenutačna dva glavna izazova za čovječanstvo: klimatske promjene i kemijska kontaminacija.
Autor/Izvor:
Nettle, a Long-Known Fiber Plant with New Perspectives
Izjava institucionalnog odbora za reviziju Nije primjenjivo. Izjava o informiranom pristanku Nije primjenjivo. Izjava o dostupnosti podataka Nije primjenjivo. Sukob interesa Autori izjavljuju da nema sukoba interesa. Bilješke Napomena izdavača: MDPI ostaje neutralan u pogledu tvrdnji o nadležnosti u objavljenim kartama i institucionalnim afilijacijama.
Reference*
*